The Microscopic Marvel of Plant Reproduction
Pollen, a ubiquitous and often overlooked component of our natural world, plays a pivotal role in the reproduction of flowering plants and conifers. Far from being mere dust, each speck of pollen is a complex and intricately designed package, carrying the genetic material necessary for fertilization. Understanding what makes pollen requires delving into the fascinating biological processes that create and disperse these microscopic powerhouses.
The Genesis of the Male Gametophyte
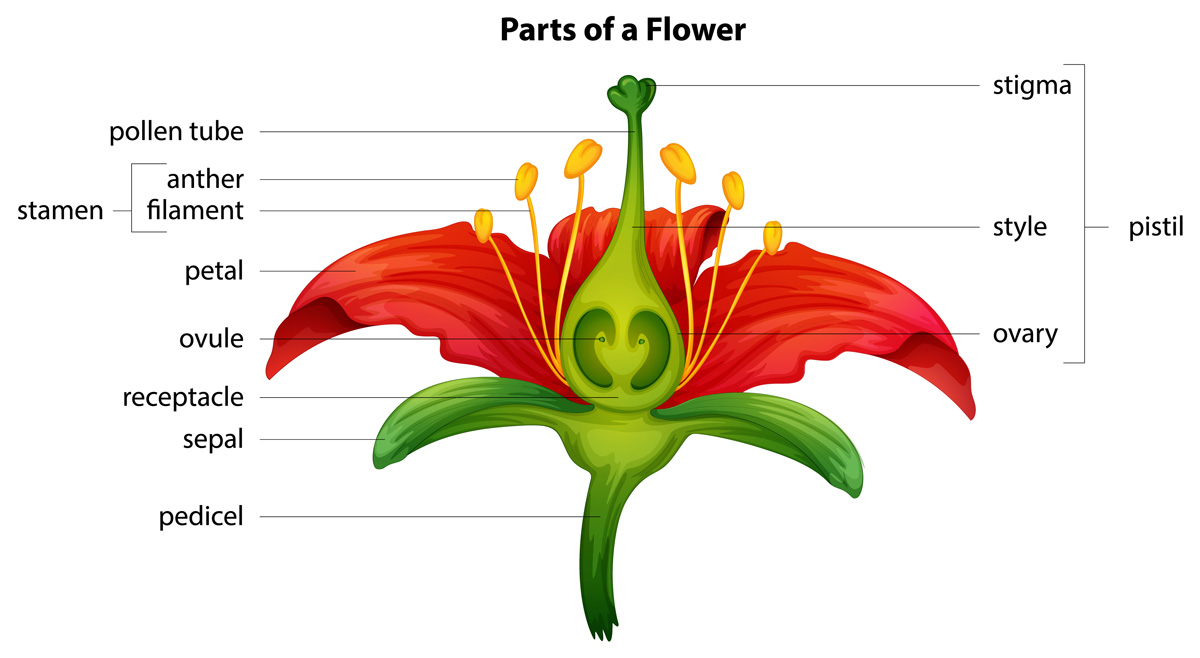
Pollen grains are, in essence, the male gametophytes of seed-bearing plants. They originate within specialized structures called anthers, which are typically found at the tips of the stamens, the male reproductive organs of a flower. Within the anther, a process known as microsporogenesis occurs, leading to the formation of pollen.
Microsporangia and Microspore Mother Cells
The anther is usually composed of two lobes, each containing one or two microsporangia, often referred to as pollen sacs. These microsporangia are the birthplaces of pollen. Inside each microsporangium are specialized diploid cells called microspore mother cells, or microsporocytes. These cells are the precursors to pollen.
Meiosis: The Foundation of Genetic Diversity
Microspore mother cells undergo meiosis, a type of cell division that reduces the chromosome number by half. This is crucial for sexual reproduction, as it ensures that when the male gamete (carried within the pollen) fuses with the female gamete (ovule), the resulting offspring will have the correct diploid number of chromosomes.
Meiosis I separates homologous chromosomes, and Meiosis II separates sister chromatids, resulting in four haploid cells called microspores. These microspores are the first haploid stage in the male gametophyte generation. Initially, these microspores are often clustered together within the microsporangium, sometimes enclosed in a shared callose wall.
Microgametogenesis: The Development into a Pollen Grain
Following meiosis, each microspore enters a phase of development known as microgametogenesis, where it differentiates into a mature pollen grain. This process involves a series of mitotic divisions and cellular changes that transform the simple microspore into a functional unit capable of delivering sperm cells.
The nucleus of the microspore divides mitotically to form two distinct nuclei: the generative nucleus and the tube nucleus (also called the vegetative nucleus). In many species, this division occurs while the microspores are still within the anther, creating a two-celled structure. In others, the generative nucleus may divide later, after the pollen grain has landed on the stigma.
The generative nucleus is responsible for producing the sperm cells, which are the actual male gametes. The tube nucleus, on the other hand, directs the growth of the pollen tube, a structure that will eventually penetrate the ovule to deliver the sperm.
The Pollen Wall: A Protective and Diagnostic Armor
One of the most remarkable aspects of pollen is its elaborate outer wall, known as the pollen wall or sporoderm. This intricate structure is composed of two primary layers: the inner intine and the outer exine. The exine, in particular, is highly sculpted and species-specific, providing crucial characteristics for plant identification.
The Intine: The Inner Layer
The intine is the inner layer of the pollen wall. It is typically composed of cellulose and pectin, similar to plant cell walls. The intine is often less sculptured than the exine and can vary in thickness. It plays a role in the hydration of the pollen grain and the emergence of the pollen tube.
The Exine: The Sculpted Exterior
The exine is the outer, and often more conspicuous, layer of the pollen wall. It is composed of sporopollenin, a highly resistant biopolymer that is exceptionally durable and resistant to degradation from chemical and physical agents, including UV radiation and decay. This resistance is key to the fossilization of pollen grains, providing invaluable insights into past plant life and environments.

The exine’s surface ornamentation is a defining feature of pollen morphology. It can be smooth, reticulate (net-like), spinulose (spiny), echinate (covered in spines), striate (lined), or exhibit a myriad of other patterns. These patterns are not random; they are genetically determined and are crucial for species identification in palynology, the study of pollen and spores.
The exine also contains specific areas called germ pores or germinal furrows. These are thin-walled regions where the exine is absent or significantly reduced. When a pollen grain germinates, the pollen tube emerges from one of these germ pores, guided by the tube nucleus. The number, size, and location of germ pores are also important taxonomic characteristics.
Pollen Release and Dispersal: The Journey to Fertilization
Once the pollen grains have matured within the anther, the final stage of their production involves their release and dispersal to facilitate fertilization. This process is highly adapted to the reproductive strategy of the plant and the environment in which it exists.
Dehiscence: Opening the Anther
The anther must open, or dehisce, to release its pollen. The mechanism of dehiscence is varied and often specific to plant families. Common methods include longitudinal slits, pores, or valves. Factors such as changes in humidity and temperature often trigger dehiscence, ensuring that pollen is released when conditions are favorable for dispersal. For example, in many species, anthers may open on dry, windy days to maximize pollen movement.
Pollination: The Transfer of Pollen
The transfer of pollen from the anther to the stigma (the receptive tip of the pistil, the female reproductive organ of a flower) is called pollination. This can occur through various agents, broadly categorized as biotic (living organisms) and abiotic (non-living factors).
Biotic Pollination
-
Entomophily (Insect Pollination): Many plants rely on insects, such as bees, butterflies, moths, and beetles, for pollination. These plants often produce showy flowers with attractive colors, scents, and nectar to lure pollinators. Pollen grains are often sticky or spiny, designed to adhere to the insect’s body. As the insect visits multiple flowers, it inadvertently transfers pollen from one to another.
-
Ornithophily (Bird Pollination): Birds, particularly hummingbirds and sunbirds, are important pollinators for certain plants. These flowers are often brightly colored (especially red or orange), tubular, and produce abundant nectar. The pollen grains are typically larger and stickier than those of wind-pollinated plants.
-
Chiropterophily (Bat Pollination): Bats, especially in tropical and subtropical regions, pollinate a variety of plants. These flowers are often large, pale or white, and open at night, emitting strong, musty, or fruity odors. They also produce copious amounts of nectar.
-
Malacophily (Snail and Slug Pollination): While less common, some plants are pollinated by snails and slugs. These flowers are often inconspicuous and may lack strong scents or bright colors.
Abiotic Pollination
-
Anemophily (Wind Pollination): A significant number of plants, including grasses, sedges, rushes, oaks, and pines, rely on wind for pollination. Wind-pollinated flowers are typically inconspicuous, lacking petals, scent, and nectar. They produce vast quantities of lightweight, smooth, and dry pollen that can be easily carried by air currents. The stigmas of these plants are often large and feathery to efficiently trap airborne pollen.
-
Hydrophily (Water Pollination): A small percentage of plants, primarily aquatic species, utilize water for pollen dispersal. This can involve pollen floating on the water’s surface or being carried beneath the surface. The pollen grains often have specialized shapes or buoyancy mechanisms.
The Role of Pollen in Fertilization
Upon successful pollination and landing on a compatible stigma, the pollen grain germinates. The tube nucleus within the pollen grain stimulates the intine to grow outward through a germ pore, forming a pollen tube. This tube grows down through the style, a stalk connecting the stigma to the ovary, and eventually reaches the ovule.
As the pollen tube elongates, the generative nucleus divides mitotically to produce two sperm cells. These sperm cells travel down the pollen tube. Upon reaching the ovule, the pollen tube penetrates the embryo sac within the ovule, and the two sperm cells are released.
In angiosperms (flowering plants), a process called double fertilization occurs. One sperm cell fuses with the egg cell to form the diploid zygote, which will develop into the embryo. The second sperm cell fuses with the central cell (containing two polar nuclei) to form the triploid primary endosperm nucleus, which will develop into the endosperm, a nutritive tissue for the developing embryo.
The intricate process of pollen formation, its robust structure, and its varied methods of dispersal highlight the remarkable evolutionary adaptations that ensure the continuation of plant life. Each pollen grain is a testament to the complexity and efficiency of nature’s reproductive strategies.